
Follow infophilosopher:
 |
|

|
|
|
Philosophers
Mortimer Adler Rogers Albritton Alexander of Aphrodisias Samuel Alexander William Alston Anaximander G.E.M.Anscombe Anselm Louise Antony Thomas Aquinas Aristotle David Armstrong Harald Atmanspacher Robert Audi Augustine J.L.Austin A.J.Ayer Alexander Bain Mark Balaguer Jeffrey Barrett William Barrett William Belsham Henri Bergson George Berkeley Isaiah Berlin Richard J. Bernstein Bernard Berofsky Robert Bishop Max Black Susanne Bobzien Emil du Bois-Reymond Hilary Bok Laurence BonJour George Boole Émile Boutroux Daniel Boyd F.H.Bradley C.D.Broad Michael Burke Jeremy Butterfield Lawrence Cahoone C.A.Campbell Joseph Keim Campbell Rudolf Carnap Carneades Nancy Cartwright Gregg Caruso Ernst Cassirer David Chalmers Roderick Chisholm Chrysippus Cicero Tom Clark Randolph Clarke Samuel Clarke Anthony Collins Antonella Corradini Diodorus Cronus Jonathan Dancy Donald Davidson Mario De Caro Democritus Daniel Dennett Jacques Derrida René Descartes Richard Double Fred Dretske John Earman Laura Waddell Ekstrom Epictetus Epicurus Austin Farrer Herbert Feigl Arthur Fine John Martin Fischer Frederic Fitch Owen Flanagan Luciano Floridi Philippa Foot Alfred Fouilleé Harry Frankfurt Richard L. Franklin Bas van Fraassen Michael Frede Gottlob Frege Peter Geach Edmund Gettier Carl Ginet Alvin Goldman Gorgias Nicholas St. John Green H.Paul Grice Ian Hacking Ishtiyaque Haji Stuart Hampshire W.F.R.Hardie Sam Harris William Hasker R.M.Hare Georg W.F. Hegel Martin Heidegger Heraclitus R.E.Hobart Thomas Hobbes David Hodgson Shadsworth Hodgson Baron d'Holbach Ted Honderich Pamela Huby David Hume Ferenc Huoranszki Frank Jackson William James Lord Kames Robert Kane Immanuel Kant Tomis Kapitan Walter Kaufmann Jaegwon Kim William King Hilary Kornblith Christine Korsgaard Saul Kripke Thomas Kuhn Andrea Lavazza James Ladyman Christoph Lehner Keith Lehrer Gottfried Leibniz Jules Lequyer Leucippus Michael Levin Joseph Levine George Henry Lewes C.I.Lewis David Lewis Peter Lipton C. Lloyd Morgan John Locke Michael Lockwood Arthur O. Lovejoy E. Jonathan Lowe John R. Lucas Lucretius Alasdair MacIntyre Ruth Barcan Marcus Tim Maudlin James Martineau Nicholas Maxwell Storrs McCall Hugh McCann Colin McGinn Michael McKenna Brian McLaughlin John McTaggart Paul E. Meehl Uwe Meixner Alfred Mele Trenton Merricks John Stuart Mill Dickinson Miller G.E.Moore Ernest Nagel Thomas Nagel Otto Neurath Friedrich Nietzsche John Norton P.H.Nowell-Smith Robert Nozick William of Ockham Timothy O'Connor Parmenides David F. Pears Charles Sanders Peirce Derk Pereboom Steven Pinker U.T.Place Plato Karl Popper Porphyry Huw Price H.A.Prichard Protagoras Hilary Putnam Willard van Orman Quine Frank Ramsey Ayn Rand Michael Rea Thomas Reid Charles Renouvier Nicholas Rescher C.W.Rietdijk Richard Rorty Josiah Royce Bertrand Russell Paul Russell Gilbert Ryle Jean-Paul Sartre Kenneth Sayre T.M.Scanlon Moritz Schlick John Duns Scotus Arthur Schopenhauer John Searle Wilfrid Sellars David Shiang Alan Sidelle Ted Sider Henry Sidgwick Walter Sinnott-Armstrong Peter Slezak J.J.C.Smart Saul Smilansky Michael Smith Baruch Spinoza L. Susan Stebbing Isabelle Stengers George F. Stout Galen Strawson Peter Strawson Eleonore Stump Francisco Suárez Richard Taylor Kevin Timpe Mark Twain Peter Unger Peter van Inwagen Manuel Vargas John Venn Kadri Vihvelin Voltaire G.H. von Wright David Foster Wallace R. Jay Wallace W.G.Ward Ted Warfield Roy Weatherford C.F. von Weizsäcker William Whewell Alfred North Whitehead David Widerker David Wiggins Bernard Williams Timothy Williamson Ludwig Wittgenstein Susan Wolf Scientists David Albert Michael Arbib Walter Baade Bernard Baars Jeffrey Bada Leslie Ballentine Marcello Barbieri Gregory Bateson Horace Barlow John S. Bell Mara Beller Charles Bennett Ludwig von Bertalanffy Susan Blackmore Margaret Boden David Bohm Niels Bohr Ludwig Boltzmann Emile Borel Max Born Satyendra Nath Bose Walther Bothe Jean Bricmont Hans Briegel Leon Brillouin Stephen Brush Henry Thomas Buckle S. H. Burbury Melvin Calvin Donald Campbell Sadi Carnot Anthony Cashmore Eric Chaisson Gregory Chaitin Jean-Pierre Changeux Rudolf Clausius Arthur Holly Compton John Conway Simon Conway-Morris Jerry Coyne John Cramer Francis Crick E. P. Culverwell Antonio Damasio Olivier Darrigol Charles Darwin Richard Dawkins Terrence Deacon Lüder Deecke Richard Dedekind Louis de Broglie Stanislas Dehaene Max Delbrück Abraham de Moivre Bernard d'Espagnat Paul Dirac Hans Driesch John Dupré John Eccles Arthur Stanley Eddington Gerald Edelman Paul Ehrenfest Manfred Eigen Albert Einstein George F. R. Ellis Hugh Everett, III Franz Exner Richard Feynman R. A. Fisher David Foster Joseph Fourier Philipp Frank Steven Frautschi Edward Fredkin Augustin-Jean Fresnel Benjamin Gal-Or Howard Gardner Lila Gatlin Michael Gazzaniga Nicholas Georgescu-Roegen GianCarlo Ghirardi J. Willard Gibbs James J. Gibson Nicolas Gisin Paul Glimcher Thomas Gold A. O. Gomes Brian Goodwin Joshua Greene Dirk ter Haar Jacques Hadamard Mark Hadley Patrick Haggard J. B. S. Haldane Stuart Hameroff Augustin Hamon Sam Harris Ralph Hartley Hyman Hartman Jeff Hawkins John-Dylan Haynes Donald Hebb Martin Heisenberg Werner Heisenberg Grete Hermann John Herschel Basil Hiley Art Hobson Jesper Hoffmeyer Don Howard John H. Jackson William Stanley Jevons Roman Jakobson E. T. Jaynes Pascual Jordan Eric Kandel Ruth E. Kastner Stuart Kauffman Martin J. Klein William R. Klemm Christof Koch Simon Kochen Hans Kornhuber Stephen Kosslyn Daniel Koshland Ladislav Kovàč Leopold Kronecker Rolf Landauer Alfred Landé Pierre-Simon Laplace Karl Lashley David Layzer Joseph LeDoux Gerald Lettvin Gilbert Lewis Benjamin Libet David Lindley Seth Lloyd Werner Loewenstein Hendrik Lorentz Josef Loschmidt Alfred Lotka Ernst Mach Donald MacKay Henry Margenau Owen Maroney David Marr Humberto Maturana James Clerk Maxwell Ernst Mayr John McCarthy Warren McCulloch N. David Mermin George Miller Stanley Miller Ulrich Mohrhoff Jacques Monod Vernon Mountcastle Emmy Noether Donald Norman Travis Norsen Alexander Oparin Abraham Pais Howard Pattee Wolfgang Pauli Massimo Pauri Wilder Penfield Roger Penrose Steven Pinker Colin Pittendrigh Walter Pitts Max Planck Susan Pockett Henri Poincaré Daniel Pollen Ilya Prigogine Hans Primas Zenon Pylyshyn Henry Quastler Adolphe Quételet Pasco Rakic Nicolas Rashevsky Lord Rayleigh Frederick Reif Jürgen Renn Giacomo Rizzolati A.A. Roback Emil Roduner Juan Roederer Jerome Rothstein David Ruelle David Rumelhart Robert Sapolsky Tilman Sauer Ferdinand de Saussure Jürgen Schmidhuber Erwin Schrödinger Aaron Schurger Sebastian Seung Thomas Sebeok Franco Selleri Claude Shannon Charles Sherrington Abner Shimony Herbert Simon Dean Keith Simonton Edmund Sinnott B. F. Skinner Lee Smolin Ray Solomonoff Roger Sperry John Stachel Henry Stapp Tom Stonier Antoine Suarez Leo Szilard Max Tegmark Teilhard de Chardin Libb Thims William Thomson (Kelvin) Richard Tolman Giulio Tononi Peter Tse Alan Turing C. S. Unnikrishnan Nico van Kampen Francisco Varela Vlatko Vedral Vladimir Vernadsky Mikhail Volkenstein Heinz von Foerster Richard von Mises John von Neumann Jakob von Uexküll C. H. Waddington James D. Watson John B. Watson Daniel Wegner Steven Weinberg Paul A. Weiss Herman Weyl John Wheeler Jeffrey Wicken Wilhelm Wien Norbert Wiener Eugene Wigner E. O. Wilson Günther Witzany Stephen Wolfram H. Dieter Zeh Semir Zeki Ernst Zermelo Wojciech Zurek Konrad Zuse Fritz Zwicky Presentations Biosemiotics Free Will Mental Causation James Symposium |
John Carew Eccles
Few scientists have had more insight into the physical workings of the mind than the great neurobiologist John Eccles. His partnership with Karl Popper produced years of conversation about the interaction between a non-material mind and the material brain. He and Popper were both dualists. His "philosophical position was diametrically opposite to those who would relegate conscious experience to a mere epiphenomenon." (Eccles, Facing Reality, 1970, p.1.)
Conversations with Popper sometimes denied any role for quantum uncertainty. That would only be chance, which could not provide moral responsibility for human actions. But Popper did produce a two-stage model for free will in 1965, under the influence of Arthur Holly Compton. Eccles wrote and lectured frequently on free will. We provide an excerpt from Facing Reality on Free Will and Creativity. There is no sign here of Popper's idea, nor of Eccles' radical hypothesis that mind used the immaterial probability wave function, based on an observation of Henry Margenau.
In this selection, Eccles has reworked the 1952 conclusion of his Waynflete lectures at Oxford - The Neurophysiological Basis of Mind, Oxford, 1953, pp.271-286.
Facing Reality, 1970, pp.118-129
From Chapter VIII, Man, Freedom, and Creativity. 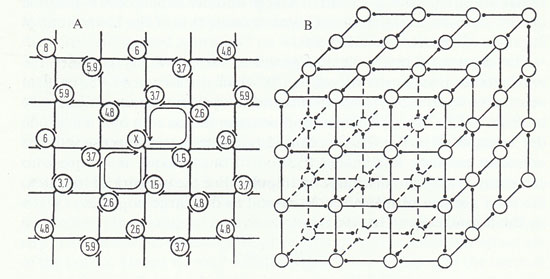
If each neurone receives and gives three synaptic contacts, a similar construction with radial symmetry is possible in three-dimensional network (Fig. 28 B). As shown in Fig. 28 these constructions would give alternating direction of transmission in the successive lines in any plane. Similarly, if each neurone gave and received n synapses, the pattern could be accommodated to an n-dimensional network.
The problem of the mode of action of the will can be simplified and sharpened by considering firstly the behavior of a single neurone in the active neuronal network of the cortex. Suppose some small "influence" were exerted at a node that would make a neurone discharge an impulse at a level of synaptic excitation which would otherwise have been just ineffective, that is, in general to raise the probability of its discharge. Such a discharged impulse would in turn have an excitatory effect on all the other nodes on which it impinges, raising the probability of their discharge, and so on. If we assume, as above, that the transmission time from node to node occupies 1 msec, then, even on the two-dimensional net of Fig. 28 A, a spread to a large number of neurones is possible in, say, 20 msec, a time that is chosen because it is at the lower limit of duration of discrete mental events.
In order to frame a precise problem, we can firstly consider the schematic neuronal networks of Fig. 28 which are assumed to be cortical neurones — both the pyramidal cells and the very numerous stellate cells. We make the postulates that at zero time a neurone (for example X in Fig. 28 A) is caused to discharge an impulse into the quiescent network and that activation of one synapse is adequate to cause any neurone to discharge an impulse. For the network of Fig. 28 A. the total number of neurones, N, caused to discharge impulses is given by the formula (SAWYER, 1951):
N = 2m2 — 2m + 2
where m is the number of nodes traversed. In 20 msec m=20, the internodal time being assumed as 1 msec; hence the number of neurones activated is 762.
On the same assumptions, but with a multi-dimensional network constructed according to the conventions of Fig. 28, the number of activated neurones, N, is given, where m is large relative to n, by the general formula (SAWYER, 1951):
N ≈ (2n/n!) m2
when m = 20 (i. e. within 20 msec) and with n = 3 (Fig. 28 B), N is of the order 104. With n = 4 and 5 respectively, N is of the order 105 and 8 x 105.
These calculations are intended merely to give some indication of the large number of cortical neurones that could be affected by a discharge originating in any one. In order to apply them to our problem of how "will" could act on the cerebral cortex, it is necessary to take into account the evidence that "will" can act on the cortical neuronal network only when a considerable part of it is at a relatively high level of excitation, i.e. we have to assume that, for "will" to be operative, large population, of cortical neurones are subjected to strong synaptic bombardment, and are stimulated thereby to discharge impulses which bombard other neurones. Under such dynamic conditions it may be conservatively estimated that, out of the hundred or more synaptic contacts made by any one neurone, at least four or five would be critically effective (when summed with synaptic bombardments by other neurones) in evoking the discharge of neurones next in series. The remainder would be ineffective because the recipient neurones would not be poised at this critical level of excitability, being either at a too low level of excitation, or at a too high level, so that the neuronal discharge occurs regardless of this additional synaptic bombardment. Thus at any instant the postulated action of the "will" on any one neurone would be effectively detected by the "critically poised neurones" on which it acts synaptically.
So long as the assumed number of critically effective synaptic excitatory actions by each neurone is kept at the low levels used in the above calculations, it is probable that the conventions of the network structures of Fig. 28 give an approximate method of allowing for all the mass of feed-back connections that occur in the closed-chain linkages of the cerebral cortex (LORENTE DE NO, 1933, 1934, 1943). Further, since the cortex is approximately 3 mm thick and the mean density of neurones 40,000 per sq. mm of surface (THOMPSON, 1899), the spread to some hundreds of thousands of neurones can be treated as spreading indefinitely in all directions without serious restriction by the sheet-like structure of the cortex. Hence we may conclude that, when a region of the cortical neuronal network is at a high level of activity, the discharge of an impulse by any one neurone will have contributed directly and indirectly to the excitation of hundreds of thousands of other neurones within the very brief time of 20 msec.
A Neurophysiological Hypothesis of Will
As a restatement of the conclusion of the preceding section we may say that in the active cerebral cortex within 20 msec the pattern of discharge of even hundreds of thousands of neurones would be modified as a result of an "influence " that initially caused the discharge of merely one neurone. But further, if we assume that this "influence" is exerted not only at one node of the active network, but also over the whole field of nodes in some sort of spatio-temporal patterning, then it will be evident that potentially the network is capable of integrating the whole aggregate of "influences" to bring about some modification of its patterned activity, that otherwise would be determined by the pattern of afferent input and its own inherent structural and functional properties. Such integration would occur over hundreds of thousands of nodes in a few milliseconds, the effects exerted on any and every node being correlated in the resultant patterned activity of the surrounding hundreds of thousands of neurones. Thus in general. the spatio-temporal pattern of activity would be determined not only by (i) the micro-structure of the neural net and its functional properties as built up by genetic and conditioning factors and (ii) the afferent input over the period of short-term memory, but also (iii) by the postulated "field of mind influence." For example, in Fig. 29 the spatio-temporal pattern determined by factors (i) and (ii) is shown diagrammatically by the shaded structure bounded by the continuous line, while a possible modification by factor (iii) is indicated by the paths outlined by broken lines at B and C. Fig. 29 can be considered as showing boundaries of multilane neuronal traffic as indicated in Figs. 10 and 12.
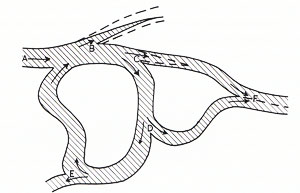
It can be claimed that no physical instrument would bear comparison with the postulated performance of the active cerebral cortex as a detector of minute "fields of influence" spread over a microscopic pattern am with temporal sequences of milliseconds. The integration, within a few milliseconds, of "influences" picked up at hundreds of thousands of nodes would be unique, particularly when it is remembered that the integration is no mere addition, but is exerted to modify in some specific way "a shifting harmony of sub-patterns" of neuronal activity, achieving expression through the modifications so produced.
Thus, the neurophysiological hypothesis is that the "will" modifies the spatiotemporal activity of the neuronal network by exerting spatiotemporal "fields of influence" that become effective through this unique detector function of the active cerebral cortex. It will be noted that this hypothesis assumes that the "will" or "mind influence" has itself some spatio-temporal patterned character in order to allow it this operative effectiveness.
The Physical Implications of the Hypothesis
When considering the manner in which mind could operate on matter, EDDINGTON (1939) discussed two hypotheses.
(i) It was postulated that mind could control the behaviour of matter within the limits imposed by HEISENBERG'S Principle of Uncertainty (cf. EDDINGTON, 1935). EDDINGTON rejected this partly because the permitted range would be exceedingly small. Presumably he was thinking of an object as large as a neurone. However, a neurophysiologist would now consider the much smaller synaptic vesicle (cf. Figs. 3, 4, 31) as the key structure on which a "mind influence" might work. The synaptic vesicle is approximately a sphere 400 A in diameter and so would have .9 mass of about 3 x 10-17 g. If, as EDDINGTON implies, the uncertainty principle is applicable to an object of this size, then it may be calculates that there is an uncertainty in the position of such an object of abou~ 50 A in I millisecond. These values are of interest since 50 A is approximately the thickness of the presynaptic membrane across which the vesicle discharges its content of specific transmitter substance.
Furthermore, as shown above, minute "influences" thus exerted or a large population of neurones would be rapidly integrated in the forn: of a changed spatiotemporal pattern of activity in the neuronal net There is thus in the active cortex a mechanism that could enormously amplify minute effects exerted on the individual synaptic vesicles, provides of course, as postulated above, these influences have some "meaningful' pattern and are not random. It is therefore possible that the permitted range of behaviour of a synaptic vesicle may be adequate to allow for the effective operation of the postulated "mind influences" on the active cerebral cortex. However, EDDINGTON rejected this hypothesis for the further reason that it involved a fundamental inconsistency. First behaviour according to chance was postulated in making a calculation of the permitted limits according to the uncertainty principle, then it was restricted or controlled by a non-chance or volitional action (the mind influence), which necessarily must be introduced if mind is to be able to take advantage of the latitude allowed by the uncertainty.
(ii) As a consequence of this rejection, EDDINGTON was led to an alternative hypothesis of a correlated behaviour of the individual particles of matter, which he assumed to occur for matter in liaison with mind The behaviour of such matter would stand in sharp contrast to the uncorrelated or random behaviour of particles that is postulated in physics and, as he stated, may be "regarded by us as something outside physics" (EDDINGTON, 1939).
Either of EDDINGTON's hypotheses could serve as the physical basis of the neurophysiological hypothesis that has here been developed for mind-brain liaison. This latter hypothesis of mind-brain liaison has the merits of relating the occasions when the mind can operate on the brain to the observed high level of neuronal activity during consciousness, and of showing how an effective action could be secured by a spatiotemporal pattern of minute "influences." If the neuronal activity of the cerebral cortex is at too low a level, then liaison between mind and brain ceases. The subject is unconscious as in sleep, anaesthesia, coma. Perception and willed action are no longer possible. Furthermore, if a large part of the cerebral cortex is in the state of the rigorous driven activity of a convulsive seizure, there is a similar failure of brain-mind liaison, which is likewise explicable by the deficiency of the sensitive detectors, the critically poised neurones.
General Discussion of Hypothesis of Free-Will
It will be sufficiently evident that the hypotheses here developed are of a fragmentary and tentative character, but it is hoped that they may be of value in further theoretical developments on mind-brain liaison. An outstanding problem for consideration would concern the postulated action of the mind in a spatiotemporal pattern, for presumably it must so act if it is to cause significant modification in patterned activity of the cortex. However, that problem would appear less formidable if there were a sufficiently rapid and detailed feed-back from brain action to mind, which in any case must be assumed for perception.
It will be objected that the essence of the hypothesis is that mind produces changes in the matter-energy system of the brain and hence must be itself in that system (cf. SCHRODINGER, 1951). But this deduction is merely based on the present hypotheses of physics. Since these postulated "mind influences" have not been detected by any existing physical instrument, they have necessarily been neglected in constructing the hypotheses of physics, as was recognized by EDDINGTON (1939). It is at least claimed that the active cerebral cortex conceivably could be a detector of such "influences" even if they existed at an intensity below that detectable by physical instruments.
The present hypotheses would offer an explanation of the high development of matter-mind traffic in the active human cerebral cortex, the development including not only continuous operation but also exquisite subtlety in transmission. Both these features would receive explanation on the basis of the interlocking, integrating, and everchanging pattern of activity formed by the numerous sensitively poised detectors (probably hundreds of millions) that exist in the cortex during states of consciousness
.
It should be pointed out that, in the discussion of the functioning of the brain in Chapters 11, III, it has initially been regarded as a "machine" operating according to the laws of physics and chemistry. In conscious states (Chapter IV) it has been shown that it could be in a state of extreme sensitivity as a detector of minute spatio-temporal fields of influence. The hypothesis is here developed that these spatiotemporal fields of influence are exerted by the mind on the brain in willed action. If one uses the expressive terminology of RYLE (1949), the "ghost" operates a "machine," not of ropes and pulleys, valves and pipes, but of microscopic spatio-temporal patterns of activity in the neuronal net woven by the synaptic connections of ten thousand million neurones, and even then only by operating on neurones that are momentarily poised close to a just threshold level of excitability. It would appear that it is the sort of machine a "ghost" could operate, if by ghost we mean in the first place an "agent" whose action has escaped detection even by the most delicate physical instruments.
But even if the hypotheses of brain-mind liaison here developed are on the right track, they are still extremely inadequate. For example, we have no concept of the nature of the mind that could exert these "ghost-like" influences. Again, the slight and irregular telepathic communications being excepted, it is not possible to answer the question: how is it that a given self is in liaison exclusively with a given brain? A further problem concerns the presumed spatiotemporal patterning of the mind. For example, is this altered, as may be operatively desirable, as the microstructure of the brain alters with developing experience and the consequent storage of memories?
2. Freedom and Creativity
It would be generally accepted that creative imagination is the most profound of human activities. It provides the illumination of a new insight or understanding. In science, creative imagination gives that revelation of a new hypothesis embracing and transcending the older hypotheses. There is an immediate aesthetic appeal in its simplicity and scope. Nevertheless it has to be subjected to rigorous criticism and experimental testing. In the most striking examples the illumination has had the suddenness of a flash, as with KEKULE and the benzene ring, DARWIN and the theory of evolution, HAMILTON and his equations. Yet with most of the great scientific hypotheses there was not this instantaneous and apparently miraculous birth of a "brain child." Rather were they developed in stages, being perfected and shaped by critical reason, as with PLANCK and the quantum theory and with EINSTEIN and the theory of relativity. Nor is the suddenness of illumination any guarantee of the validity of a hypothesis. I have had only one such sudden illumination — the so-called Golgi-cell hypothesis of inhibition (BROOKS and ECCLES, 1947) — and some years later it was proved false (ECCLES, 1953)!
If I reflect on the happenings during a scientific investigation, I find that there is incessant "traffic" between my conscious experiences and the objects and events in the external world. For example, from the framework of scientific knowledge I derive some ideas about what I should observe under certain experimental conditions. I plan these conditions and then by means of controlled movements proceed to actualize these conditions. My observations or conscious experiences of the ensuing results are correlated and evaluated in rational and critical thought against my original ideas, and further experiments are planned and executed, and so on. The consequence is that my scientific ideas or hypotheses are enriched, or changed or falsified. My scientific activity is thus seen to be essentially an affair of my rational and conceptual thought together with my exercise of willed movement and my sensory perception.
Before attempting to picture the brain activities that underlie creative imagination, it is important to realize that such illuminations, whether flash-like or with a more gradual and controlled development, come only to minds that have been prepared by the assimilation and critical evaluation of the knowledge in that particular field. One can deliberately seek to experience some new imaginative insight by pouring into one's mind hypotheses and the related experiments and then relax to give opportunity for the subconscious processes that may lead to the illumination in consciousness of a new insight. Such illuminations are often fragmentary and require conscious modification, or so erroneous as to invite immediate rejection by critical reason. Nevertheless, they all give evidence of the creativeness of the subconscious mind.
It may now be asked: what kind of activity is occurring in the brain during this creative activity of the subconscious mind, and how eventually does this creative activity flash into consciousness? Let us consider firstly the pre-requisites for such cerebral action. The wealth of stored memories and critical evaluations implies that in the neuronal network there is an enormous development of complex highly specialized engrams (cf Chapter III) whose permanency derives from the postulated increases in synaptic efficacy. We may say that these "plastic" patterns give the know-how of the brain. We become expert in some fields of knowledge by virtue of the wealth and sublety of engrams that may extend over the greater part of the cortex. When pondering deeply on some problem in this field there must be an unimaginably complex and vivid interplay in the activated patterns. One can speculate further that some failure in the synthesis of these patterns or some conflict in their inter-relationship is the neuronal counterpart of a problem that clamours for solution.
Such are the pre-requisites leading to creative insight. We may surmise that the "subconscious operation of the mind" is dependent on the continued intense interplay of these patterns of neuronal activation. We have seen (Chapter III) that on repeated activation of any neuronal pattern there tends to be a progressive change in the basic plastic pattern or engram, particularly on account of interactions with other patterns. Thus we can expect that new emergent patterns will arise during the subconscious operation of the mind. Should an emergent pattern have an organization that combines and transcends the existent patterns we may expect some resonant-like intensification of activity in the cortex, which would bring the new transcendent pattern to conscious attention, where it would appear as a bright new idea born of creative imagination.
Then begins the process of conscious criticism and evaluation, which seeks to discover flaws in the new idea, and also to discover if it is consistent with the existing knowledge. This done, there comes the crucial stage of the design and carrying out of experiments that test predictions derived from the new idea. We may say that a creative imagination is particularly fruitful if it develops new hypotheses that are remarkable for their generality and for the manner in which they stand up to crucial experimental tests.
Finally, we may ask: what are the characteristics of a brain that exhibits remarkable power of creative imagination? In attempting an answer we are more than ever in the realm of speculation, but certain general statements can be made, though their inadequacy is all too apparent. There must, firstly, be an adequate number of neurones, and. more importantly, there should be a wealth of synaptic connection between them, so that there is, as it were, the structural basis for an immense range of patterns of activity. It is here that the inadequacy of explanation is so evident. There is but a poor correlation between brain size and intelligence, but in this assessment one is assuming a proportionality of brain size and neurone population. Furthermore, a chimpanzee brain may have a neurone population as high as 70% of a human brain, yet it displays almost no creative imagination. Secondly, there should be a particular sensitivity of the synapses to increase their function with usage (cf. Chapter 111) so that memory patterns or engrams are readily formed and are enduring. Both of these properties will ensure that eventually there is built up in the brain an immense wealth of engrams of highly specific character. If added to this there is a peculiar potency for unresting activity in these engrams so that the spatiotemporal patterns are continually being woven in most complex and interacting forms, the stage is set for the deliverance of a "brain child" that is sired, as we say, by creative imagination.
Three Worlds
I fully accept the recent philosophical achievements of Sir Karl Popper with his concept of three worlds.
World 1 is the world of physical objects and states. It comprises the whole cosmos of matter and energy, all of biology including human brains, and all artifacts that man has made for coding information, as for example, the paper and ink of books or the material base of works of art. World 1 is the total world of the materialists. They recognise nothing else. All else is fantasy.
World 2 is the world of states of consciousness and subjective knowledge of all kinds. The totality of our perceptions comes in this world. But there are several levels. In agreement with Polten, I tend to recognise three kinds of levels of World 2, as indicated in Fig. 6-2, but it may be more correct to think of it as a spectrum.
"The first level (outer sense) would be the ordinary perceptions provided by all our sense organs, hearing and touch and sight and smell and pain. All of these perceptions are in World 2, of course: vision with light and colour; sound with music and harmony; touch with all its qualities and vibration; the range of odours and tastes, and so on. These qualities do not exist in World 1, where correspondingly there are but electromagnetic waves, pressure waves in the atmosphere, material objects, and chemical substances.
In addition there is a level of inner sense, which is the world of more subtle perceptions. It is the world of your emotions, of your feelings of joy and sadness and fear and anger and so on. It includes all your memory, and all your imaginings and planning into the future. In fact there is a whole range of levels which could be described at length. All the subtle experiences of the human person are in this inner sensory world. It is all private to you but you can reveal it in linguistic expression, and by gestures of all levels of subtlety.
Finally, at the core of World 2 there is the self or pure ego, which is the basis of our unity as an experiencing being throughout our whole lifetime.
This World 2 is our primary reality. Our conscious experiences are the basis of our knowledge of World 1, which is thus a world of secondary reality, a derivative world. Whenever I am doing a scientific experiment, for example, I have to plan it cognitively, all in my thoughts, and then consciously carry out my plan of action in the experiment. Finally I have to look at the results and evaluate them in thought. For example, I have to see the traces of the oscilloscope and their photographic records or hear the signals on the loudspeaker. The various signals from the recording equipment have to be received by my sense organs, transmitted to my brain, and so to my consciousness, then appropriately measured and compared before I can begin to think about the significance of the experimental results. We are all the time, in every action we do, incessantly playing backwards and forwards between World 1 and World 2.
And what is World 3? It is the whole world of culture. It is the world that was created by man and that reciprocally made man. This is my message in which I follow Popper unreservedly. The whole of language is here. All our means of communication, all our intellectual efforts coded in books, coded in the artistic and technological treasures in the museums, coded in every artefact left by man from primitive times—this is World 3 right up to the present time. It is the world of civilisation and culture. Education is the means whereby each human being is brought into relation with World 3. In this manner he becomes immersed in it throughout life, participating in the heritage of mankind and so becoming fully human. World 3 is the world that uniquely relates to man. It is the world which is completely unknown to animals. They are blind to all of World 3. I say that without any reservations.
Eccles and Popper Worlds are fully consistent with the three worlds of Information Philosophy.
For Teachers
For Scholars
|



